Data
The data will be made available through the Nordicana D data website of University Laval which attributes DOIs to data sets, facilitating their citation.
First results
1-
Impact of vegetation cover on the thermal regime of the ground, Umiujaq
Umiujaq is a transition zone where within a radius of 2 km coexist, a lichen cover (Cladonia stellaris), low dwarf birch (Betula glandulosa, 30 cm), tall dwarf birch (1 m) and lastlyblack spruce stands (Picea mariana), which are in fact relics of a warmer past A bit further open forest is present, where black spruce dominates (Photos 1 and 2).

shrub tundra at the top of the valley. A relic spruce massif is visible on the left.

Photo 2 - Umiujaq site showing instruments and types of vegetation cover -
forest tundra at the bottom of the valley.
Temperature sensors have been installed at various depths in the ground in 2012. Figure 1a illustrates the impact of vegetation cover on soil temperature at 50 cm. It is clear that the higher and denser the vegetation, the more the gound is protected from winter cooling. This is explained first by the snow depth, which increases from 80 cm on lichen to 250 cm in forest tundra, but also by soil moisture (Figure 1b) which increases with plant growth, delaying or even preventing freezing of the soil due to the latent heat of freezing, longer to evacuate. In fact, permafrost has disappeared from areas where erect vegetation has appeared and only exists in some areas of low vegetation.
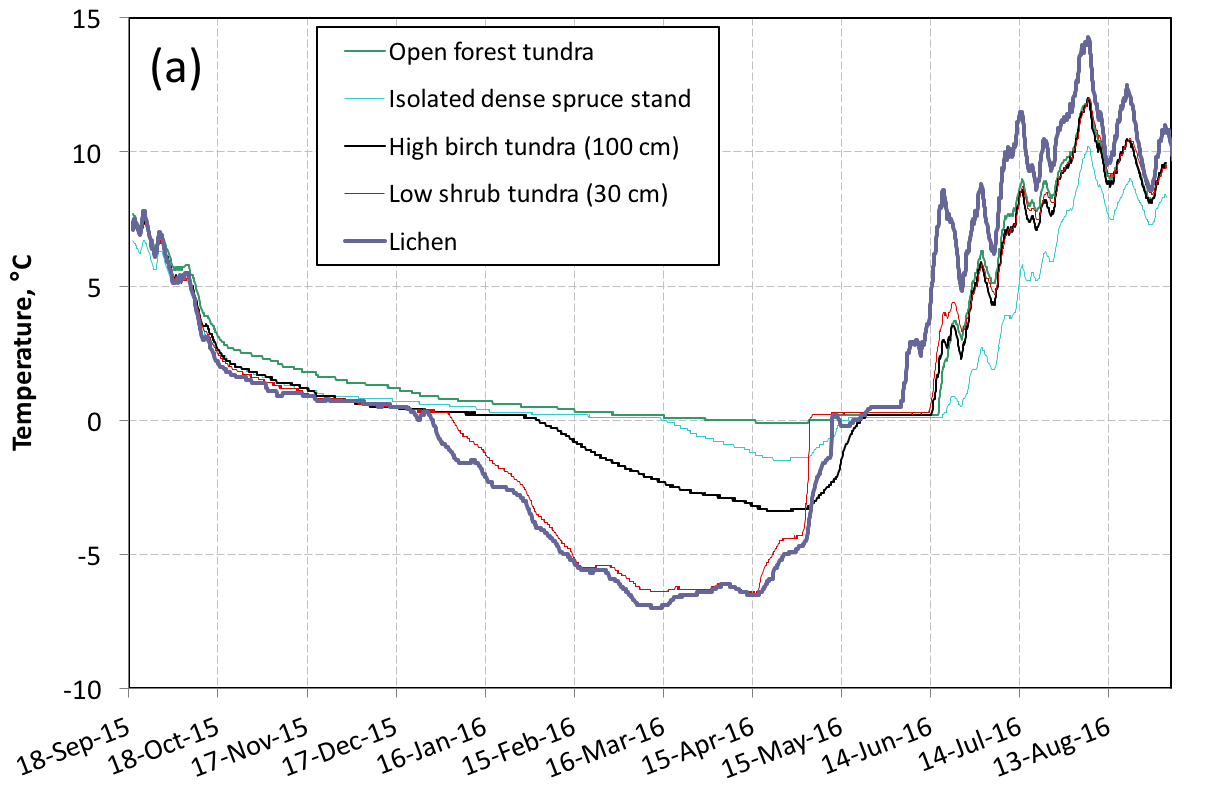
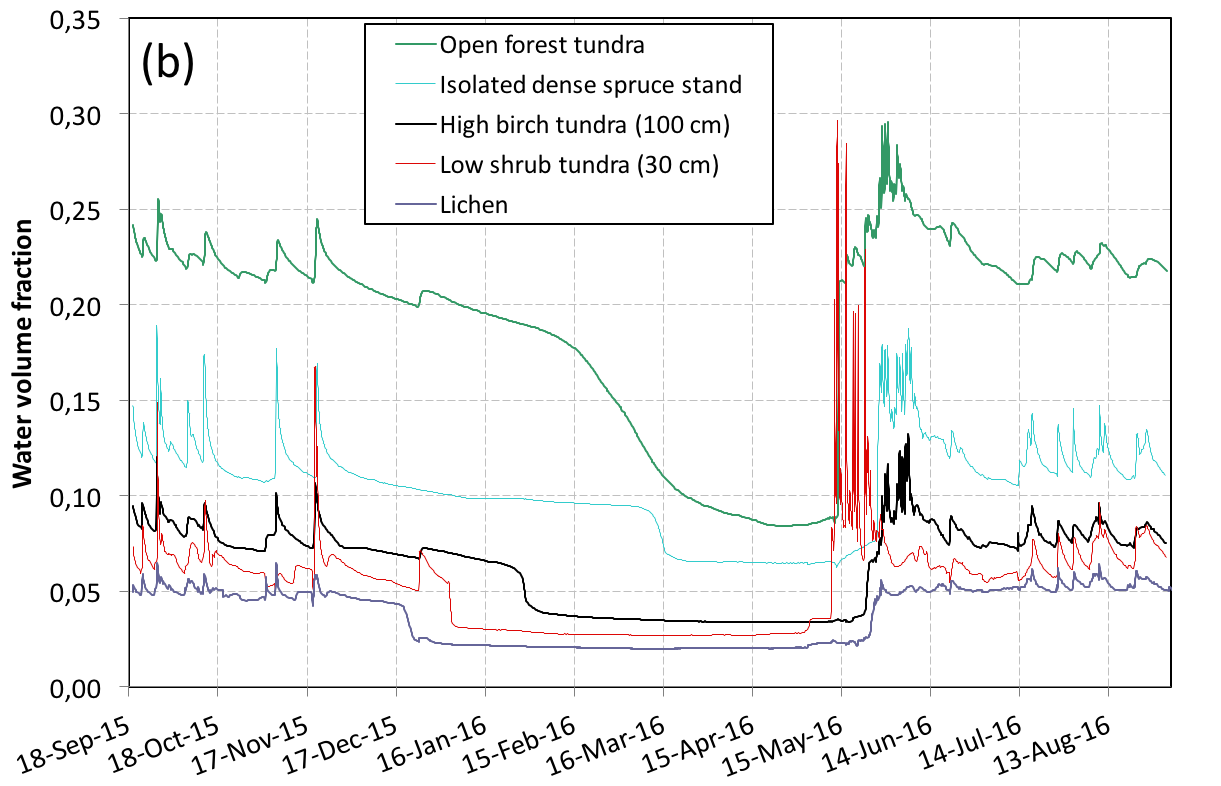
Summer data illustrate another impact of vegetation on the ground energy budget. The lichen does not produce shading on the ground and forms a layer with limited thermal insulation, so that the lichen soil is the hottest in summer. The dense spruce stands produce almost total shade and the thick moss layer at their base effectively insulates the ground, limiting warming. With regards to the annual budget, low shrubs are the most effective at keeping the ground cool: they have no noticeable effect on snow accumulation, because the snow depth exceeds their own height, while providing shade that limits summer warming. These data illustrate the complexity of snow-vegetation-permafrost interactions, and the challenge of modeling them. Indeed, many processes highlighted here (and others not yet discussed) are not taken into account by the most sophisticated land surface models. For example, these models do not differentiate moss and lichen, despite their very different thermal and hydrological properties.
2- Willow growth on Bylot Island, High Arctic
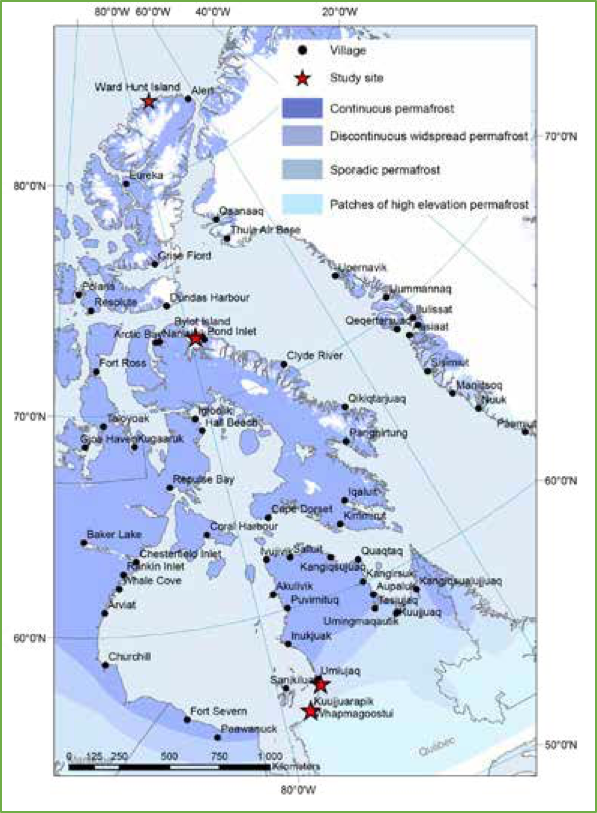
Bylot Island, north of Baffin Island (Figure 2), is a very cold site in Canada's high Arctic (mean annual temperature -14.5 °C) with continuous and thick permafrost (> 400 m).
The vegetation is mostly herbaceous, but there are a few sites with erect willows (Photo 3): Salix richardsonii up to 40 cm and Salix arctica, often prostrate but occasionally erect with a height of 15 cm.

Willows, being quite low, they only moderately favor snow accumulation, but they modify snow physical properties. Our winter campaigns have quantified the impact of the presence of Salix richardsonii on the physical properties of snow, and we present in Figure 3 the results concerning density and thermal conductivity.
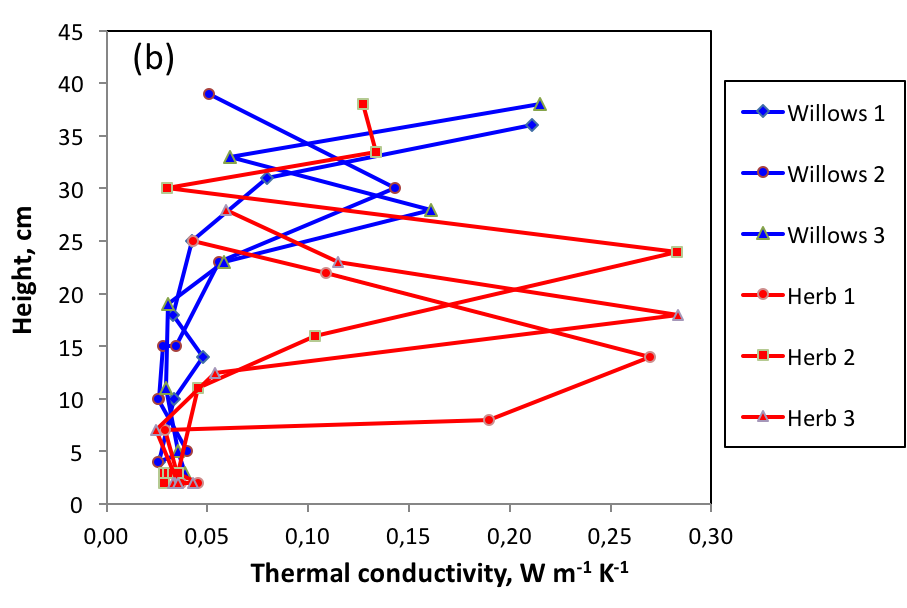
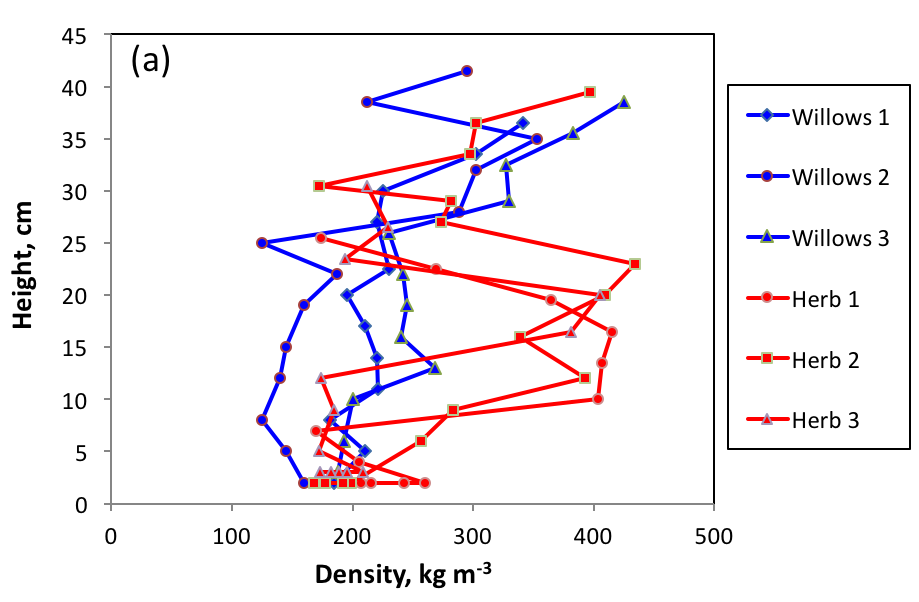
Figure 3 - Density (a) and thermal conductivity (b) of snow on herbaceous tundra and in the
willows at Bylot Island in May 2015.
Willows prevent the compaction of snow, which is therefore of low density, whereas on the herbaceous tundra the action of the wind forms much denser wind slabs (Figure 3a). These conditions favor the transformation of snow into depth hoar over almost the entire thickness of the snowpack in the willows. This type of snow has excellent thermal insulation properties (Figure 3b), which limits the winter cooling of the soil. These processes were modeled using the Crocus snow model and the ISBA land surface model. The results (Figure 4) indicate that winter soil temperature that can be up to 13 °C higher under willows.
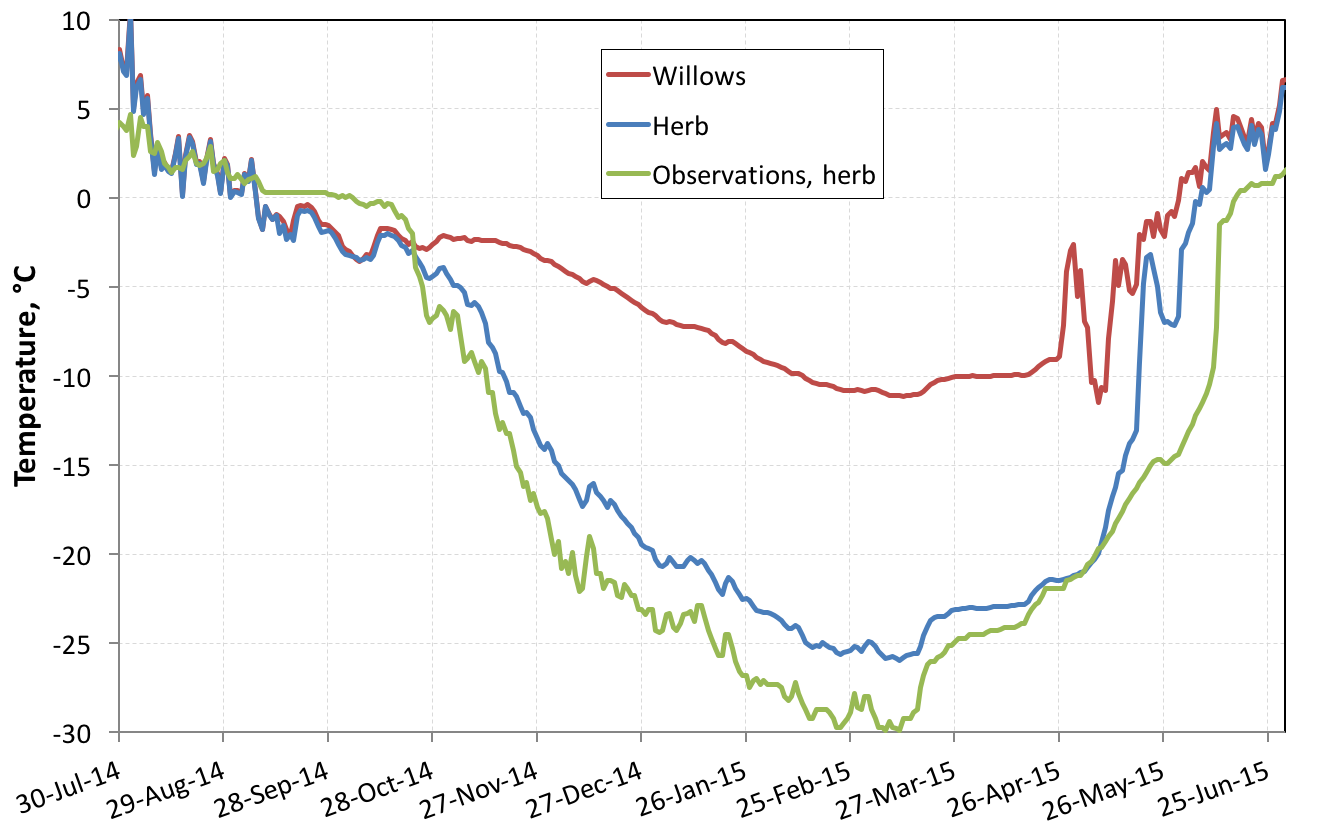
Figure 4 - Modeling soil temperature at 5 cm depth for two types of plant cover: willows and herb. Measurements obtained under herb are also reported.
The validation of this simulation was delayed because ground temperature sensors under the willows were chewed by foxes. These results show that the presence of shrubs can significantly limit ground winter cooling, contributing to permafrost warming. However, a definitive and general conclusion about the role of shrubs is doubtless premature. First of all, conditions are different each year and it is possible that different weather conditions can lead to different results. For example, shrubs absorb solar radiation and therefore warm the snow in the fall, which could lead to the formation of dense and conductive crusts of refrozen snow. Although this process has not yet been observed at Bylot, it could lead to more severe winter cooling under the willows. Multi-year monitoring is therefore essential to detect all the processes involved in vegetation-snow-permafrost interactions.
WIND SLABS
|
Second, modeling these processes is currently insufficiently developed. In addition to the details of the impacts of the various types of vegetation mentioned above, Arctic snow modeling is unsatisfactory, in particular because it does not correctly reproduce depth hoar formation. This results in a poor simulation of ground freezing in the fall and its thaw in spring, as shown in Figure 4 .
Another example is the difficulty of simulating the impact of willows on the optical properties of snow, an aspect where experimental data is sorely lacking.
3- Birch growth in Umiujaq and the ground carbon budget
Soils of the Tasiapik Valley near Umiujaq are sandy. They are emerged beaches, raised more than 150 m by isostatic rebound following the melting of the Laurentide ice sheet 15,000 to 6,000 years ago. These sandy soils are very poor in organic matter, <1 kg m-2. Under lichen areas, litter is very thin, and becomes thicker as birches grow (Photos 4 and 5). We analyzed vertical soil carbon profiles for various types of vegetation cover. At the same time, we dated the birches by dendrochronology. This was done by sampling the root-stem junction of the shrub. The objective was to quantify the rate of soil carbon accumulation due to birch growth.


Photo 4 and 5 - Soil profiles showing litter differences under lichens (Cladonia stellaris) and birches (Betula glandulosa).
We examined the correlation between the amount of carbon in the soil and the age of the birches. The correlation was low (R2 = 0.17) because the growth rate of birchshrubs, evaluated by growth ring thickness (Photo 6), is very variable depending on the site. For example, shrubs in hollows, where snow accumulation is favored, grow faster than others located on hilltops.

Photo 6 - Thin section of dwarf birch allowing the determination of its age by counting growth rings. Note that some rings have a thickness <10.
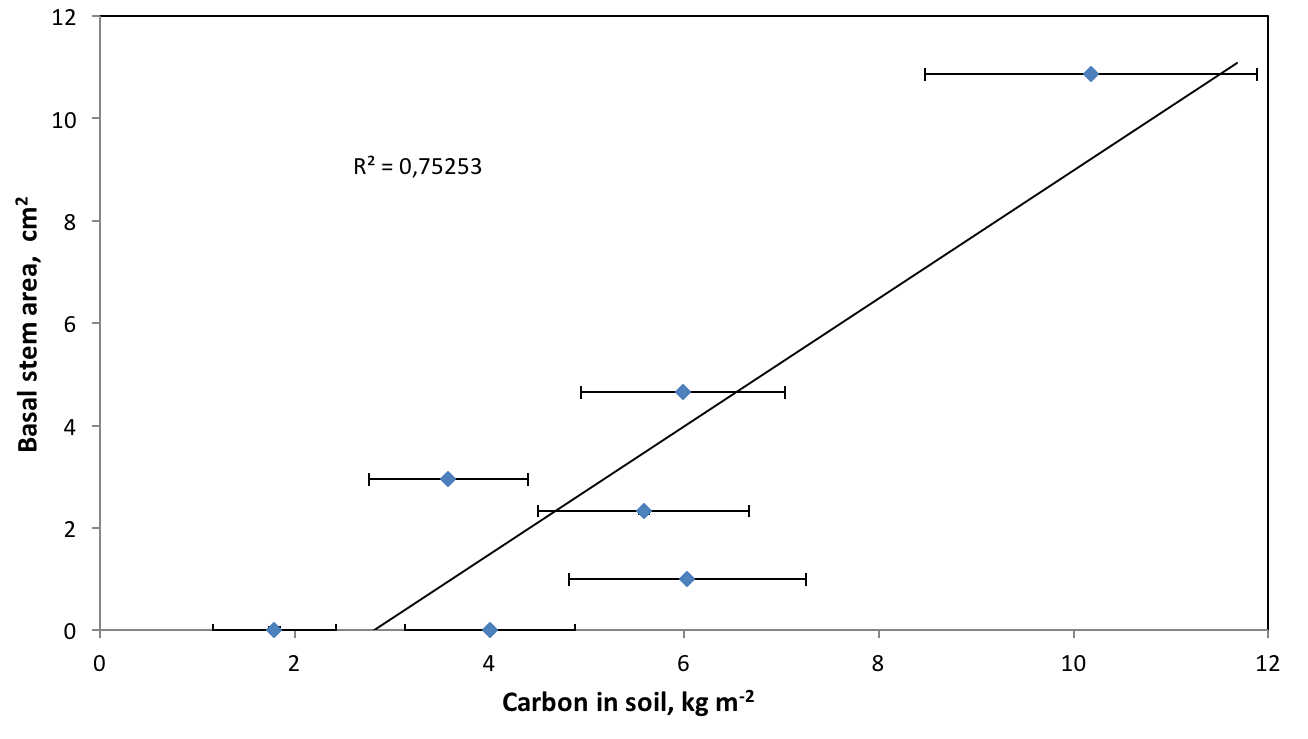
Stem diameter, an indicator of biomass accumulated by birches, was found to be much better correlated with soil carbon (Figure 5).
This simply means that the amount of carbon accumulated in the soil is approximately proportional to the birch biomass. However, the annual accumulation rate strongly depends on the site, which determines the growth conditions of the birches. The average growth rate on the 5 birch sites studied is 0.19 kg C m-2 year-1.
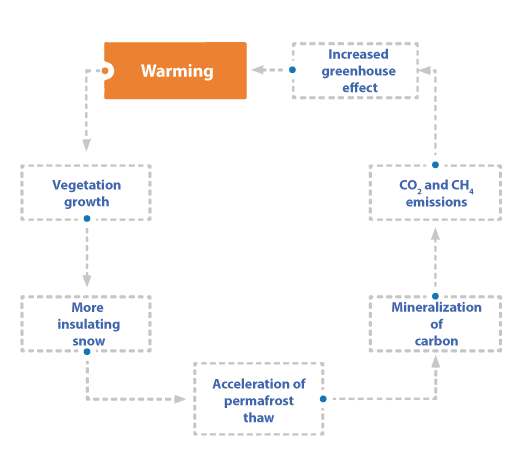
CONCLUSION
Processes affecting the energy and carbon budget of permafrost are complex and many are not described in today's most sophisticated models, which severely limits our ability to project the evolution of permafrost. The two examples of snow-vegetation interactions detailed here suggest that shrub growth on tundra leads to ground winter warming. However, summer processes such as shading by shrubs or the growth of an insulating moss layer beneath them can modulate winter effects. A wide variety of winter weather conditions, many of which have not yet been observed, could also alter the properties of the snow, leading to a different conclusion. Long-term monitoring is therefore essential for reliable quantification and modeling of the feedbacks in Figure 6. With regard to the carbon budget of permafrost, quantifying the impact of shrub growth has been difficult in the past, because separating this term from the microbial respiration of old carbon can be a source of uncertainty. In Umiujaq, where the latter process is weak, our preliminary work indicates that shrubs are a significant carbon sink, but its precise determination requires broader sampling.